

Next Lesson - The Placenta
Core
Introduction
Fetal physiology is unusual in that it is preparation for a life that has not yet started. The fetus must support its own metabolism, growth and development inside a tightly regulated maternal environment, while building the organs; lungs, kidneys, gut, immune system; that will only function fully after birth. Understanding the trade-offs the fetus makes; oxygen-poor blood handled by oxygen-greedy haemoglobin, lungs filled with fluid and bypassed by the circulation, kidneys producing urine that contributes to amniotic fluid rather than to waste clearance; explains both healthy fetal growth and most of the early neonatal emergencies.
This article focuses on the physiology of the term fetus and the changes at birth. The architecture of the placenta is covered next in The Placenta; embryology of organ formation is in Development of the Reproductive System and the system-specific embryology articles in Embryology; the maternal cardiovascular and respiratory adaptations in pregnancy are in Physiology of the Mother.
Maternal-Fetal Exchange in Outline
The placenta is the fetus's lungs, gut, kidney and liver. Practical points relevant to fetal physiology:
- Oxygen exchange. Maternal arterial blood entering the placenta has a PO2 of about 95 mmHg, but the blood emerging in the umbilical vein has a PO2 of only 30-35 mmHg. Fetal physiology functions at this oxygen tension throughout intrauterine life. The fetal-to-placental difference is offset by HbF's higher oxygen affinity, the higher haemoglobin concentration, and the higher cardiac output per kilogram in the fetus.
- Glucose crosses the placenta down its concentration gradient via GLUT1 transporters. Glucose is the principal fetal fuel.
- Amino acids are actively transported across the placenta against a concentration gradient, by Na+-coupled cotransporters.
- Free fatty acids cross by passive diffusion, more slowly.
- CO2 diffuses readily; the fetal PCO2 is slightly higher than maternal.
- Waste: urea, creatinine, bilirubin: is cleared via maternal liver and kidney.
Fetal Cardiovascular Physiology
The Three Shunts
The fetal circulation looks complicated at first glance because it must achieve two goals simultaneously: deliver oxygenated placental blood to the body, and bypass the non-functioning lungs and liver. It does both with three shunts; one venous bypass of the liver, and two right-to-left cardiac shunts that bypass the lungs. All three close after birth.
- Ductus venosus. Connects the umbilical vein (carrying the most oxygenated blood in the fetus) to the inferior vena cava, bypassing most of the liver. About 50% of umbilical venous blood takes this short cut; the rest perfuses the liver. Because the fetal hepatic metabolism is not yet running at full capacity, the liver is bypassed deliberately to deliver the freshest blood to the heart.
- Foramen ovale. An opening between the right and left atria, with a flap (the septum primum) acting as a one-way valve. Inferior vena caval blood arriving at the right atrium is preferentially streamed across the foramen ovale into the left atrium, again bypassing the lungs.
- Ductus arteriosus. Connects the pulmonary artery directly to the descending aorta distal to the origin of the great vessels supplying the head and upper limbs. Because pulmonary vascular resistance in the fetus is high (the lungs are unexpanded and hypoxic), most of the right ventricular output preferentially takes this route into the systemic circulation, bypassing the lungs once again.
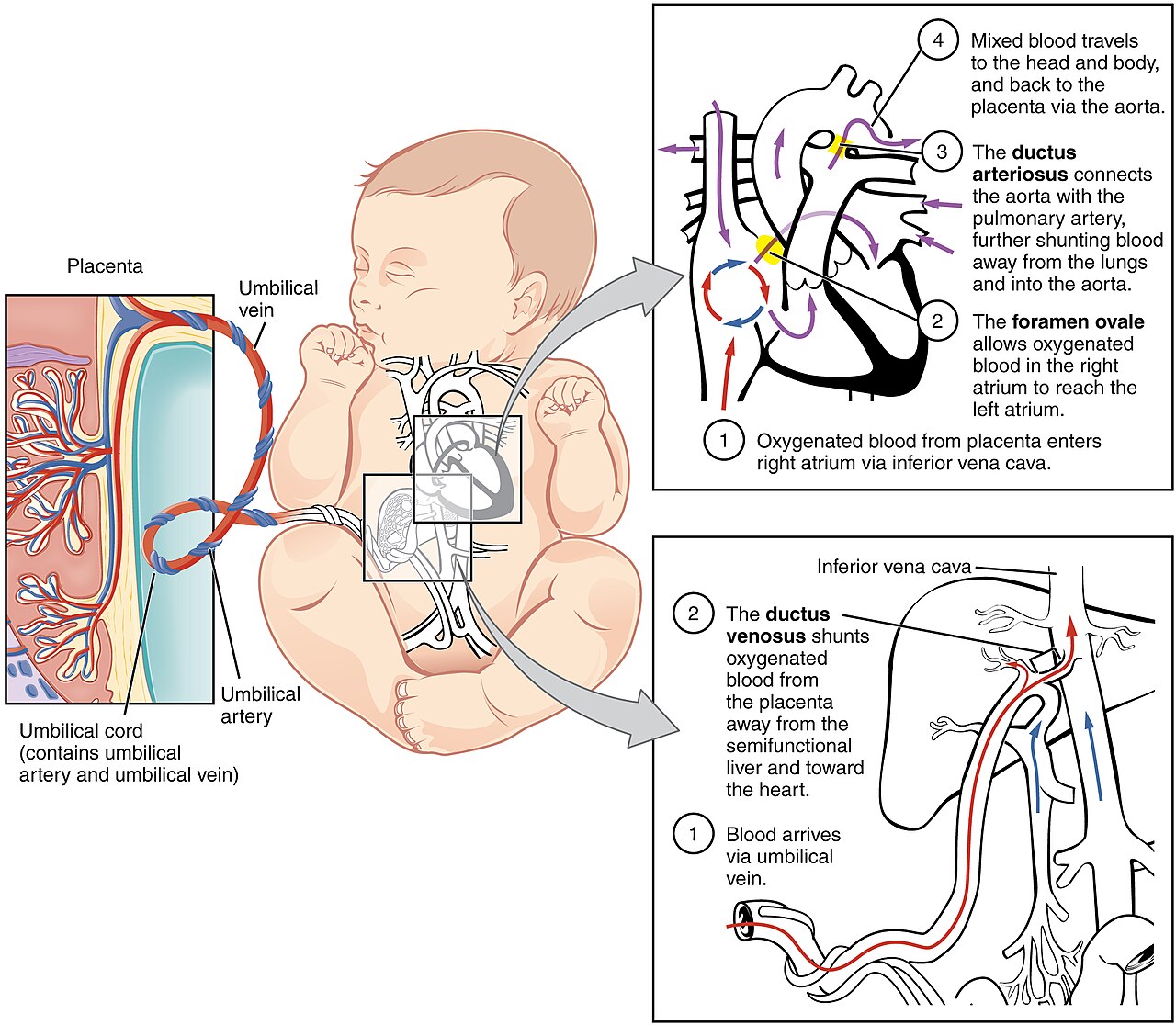
Diagram: the fetal circulation, showing the three right-to-left shunts that bypass the liver (ductus venosus) and lungs (foramen ovale and ductus arteriosus). The colour gradient highlights how the most oxygenated blood is streamed preferentially to the brain and myocardium.
Creative commons source by OpenStax College, CC BY 3.0.
Circulation and Streaming
The path of fetal blood, traced from the placenta:
- Oxygenated blood enters the fetus via the single umbilical vein.
- Half passes through the ductus venosus into the inferior vena cava, where it joins less-oxygenated blood from the lower body. Anatomical streaming inside the right atrium directs this oxygen-rich stream preferentially across the foramen ovale into the left atrium and on to the left ventricle.
- From the left ventricle, blood is ejected into the ascending aorta and supplies the brain and myocardium: the two organs that need the highest oxygen tension.
- Meanwhile, less-oxygenated blood returning from the upper body via the superior vena cava passes through the right atrium into the right ventricle and then into the pulmonary artery.
- Most of this passes through the ductus arteriosus into the descending aorta. It is now mixed with the residual oxygenated blood from the left ventricle, supplies the lower body, and returns to the placenta via the two umbilical arteries.
Three points to lock in:
- The umbilical vein carries oxygenated blood: the opposite of every other vein in the body.
- The two ventricles work in parallel, not in series; they share a single combined cardiac output, with the right ventricle contributing about two-thirds of it.
- Streaming is functional rather than absolute: there is mixing at every step, so even the ‘best’ blood reaching the brain has a PO2 of only about 25-30 mmHg, much lower than postnatal arterial values.
Fetal Haemoglobin
The fetus copes with this low-oxygen environment by using fetal haemoglobin (HbF, α2γ2) instead of adult HbA (α2β2). The two γ chains differ from β chains in their interaction with 2,3-bisphosphoglycerate (2,3-BPG): HbF binds 2,3-BPG poorly. Because 2,3-BPG normally stabilises the deoxygenated (T) state of haemoglobin, HbF in its absence binds oxygen more avidly. The oxygen dissociation curve is therefore shifted to the left compared with HbA, for any given PO2, fetal blood holds more oxygen.
HbF predominates from the second trimester until shortly after birth. The switch from γ-chain to β-chain synthesis begins late in the third trimester and is largely complete by 6 months postnatally. The principles of oxygen transport are explained in Protein Function in Oxygen Transport; the genetic basis of haemoglobinopathies is in Haemoglobinopathies.
Fetal blood also carries a higher haemoglobin concentration (around 17 g/dL at term) and a higher cardiac output per kilogram, both of which add to oxygen delivery despite the low PO2.
Fetal Respiratory Physiology
Stages of Lung Development
The lungs develop in five overlapping stages, all of which are commonly tested:
- Embryonic (weeks 3-7): lung bud arises from the foregut; main bronchi formed.
- Pseudoglandular (weeks 5-17): the conducting airway tree is laid down, looking like a branching gland under the microscope. No respiratory exchange surface yet.
- Canalicular (weeks 16-26): respiratory bronchioles develop and become vascularised. From around 24 weeks, type II pneumocytes start producing surfactant. This explains the lower limit of viability historically being 24 weeks.
- Saccular (weeks 24-38): terminal sacs form and thin out their walls.
- Alveolar (week 36 to ~8 years postnatally): mature alveoli appear and multiply. Most alveolar formation is postnatal.
Surfactant
Surfactant is a phospholipid-protein mixture that lowers alveolar surface tension, preventing alveolar collapse at end-expiration. It is produced by type II pneumocytes from approximately 24 weeks gestation, with production rising rapidly through the third trimester. The principal phospholipid is dipalmitoylphosphatidylcholine (DPPC); the surfactant proteins SP-A through SP-D contribute to organisation and host defence.
Without sufficient surfactant, alveoli collapse on each expiration and the work of breathing becomes overwhelming; the picture of respiratory distress syndrome (RDS) of the newborn. Two interventions reduce its incidence and severity:
- Antenatal maternal corticosteroids (typically betamethasone or dexamethasone) given to women at risk of preterm delivery, accelerating type II pneumocyte maturation and surfactant production.
- Exogenous surfactant replacement: animal-derived surfactant given to the preterm neonate after birth to support pulmonary function.
Fetal Breathing Movements
The fetus performs fetal breathing movements from the second trimester. The lungs are filled with lung fluid, not air, and gas exchange does not occur, but the diaphragm contracts rhythmically. These movements are essential for normal lung development, particularly the stretch-driven proliferation of pneumocytes. Conditions that suppress fetal breathing or restrict thoracic expansion (oligohydramnios, congenital diaphragmatic hernia, neuromuscular disease) cause pulmonary hypoplasia.
The lung fluid produced during gestation is mostly cleared in two phases at birth: a fall in production triggered by the rise in adrenaline during labour, and active sodium absorption through ENaC channels in the epithelium driven by the same adrenergic surge. Mechanical squeezing of the chest during vaginal delivery contributes to the final clearance; one of the reasons elective caesarean delivery (without labour) is associated with transient tachypnoea of the newborn.
Fetal Renal and Gastrointestinal Physiology
The fetal kidneys begin producing urine from about 10 weeks. By the third trimester, fetal urine is the principal contributor to amniotic fluid volume. The fetus does not need its kidneys for waste clearance: the placenta does this, but adequate fetal urinary output is essential to maintain amniotic fluid volume, which in turn is essential for normal lung development and protection from cord compression.
This explains the link between renal anomalies and lung problems in the neonate. Bilateral renal agenesis (Potter sequence) produces severe oligohydramnios, pulmonary hypoplasia, and a characteristic facial appearance. Posterior urethral valves and bilateral severe hydronephrosis can produce similar picture by obstructive renal failure. Conversely, fetal anuria is one of the reasons antenatal ACE inhibitors are contraindicated; they shut off fetal renal blood flow.
The fetal gastrointestinal tract develops motility from mid-gestation. The fetus swallows amniotic fluid; this is essential for normal gut development. Conditions that impair fetal swallowing (anencephaly, gross neurological abnormality, oesophageal atresia) cause polyhydramnios. The fetal bowel accumulates meconium: a sticky greenish-black mixture of swallowed amniotic-fluid debris, intestinal cells, bile and gut-derived enzymes. Meconium is normally retained until birth. Its release into amniotic fluid (meconium-stained liquor) can indicate fetal compromise, but at term and post-term it also reflects gastrointestinal maturity and is not in itself specific for stress. Aspirated meconium at delivery is the substrate for meconium aspiration syndrome.
Fetal Endocrine and Metabolic Physiology
The fetus has a more independent endocrine system than is sometimes appreciated:
- Thyroid hormone. Maternal T4 crosses the placenta in small but functionally important amounts, particularly in the first trimester before the fetal thyroid is producing its own. Severe maternal hypothyroidism in early pregnancy, or untreated congenital hypothyroidism in the neonate; can cause irreversible neurological damage; this is the rationale for the Guthrie heel-prick screening test in the first week of life. See The Thyroid Gland.
- Cortisol. A surge of fetal cortisol in the days before labour matures multiple organ systems, including the lungs (boosting surfactant production), the gut (boosting absorptive capacity) and the liver (boosting glucose-6-phosphatase activity, allowing glycogenolysis). The same surge contributes to the initiation of labour. Antenatal exogenous corticosteroids in threatened preterm delivery exploit this system.
- Insulin. The fetal pancreas produces insulin from about 12 weeks. Maternal insulin does not cross the placenta, but maternal glucose does. In maternal diabetes, fetal hyperglycaemia drives fetal hyperinsulinaemia, which in turn drives macrosomia and increases the risk of neonatal hypoglycaemia in the first hours of life when maternal glucose supply is suddenly withdrawn.
- Calcium handling is dominated by active transplacental transport against a gradient. Fetal serum calcium and phosphate are higher than maternal. After birth this support is lost and neonatal calcium falls; transient neonatal hypocalcaemia in the first 48 hours is common, particularly in preterm or stressed infants. See Calcium Metabolism.
- Glucose metabolism. The fetus relies on continuous maternal glucose supply, with glycogen stored in liver and skeletal muscle and brown adipose tissue laid down in late pregnancy. The brown adipose tissue is essential for postnatal thermogenesis (see below). Fetal hepatic gluconeogenesis is induced rapidly after birth by the cortisol surge and falling insulin.
Fetal Immune Physiology
The fetus is an immunological compromise. It expresses paternal antigens and could in principle be rejected by the maternal immune system; instead, both maternal and fetal immune systems develop tolerance through specialised mechanisms at the placental interface (low MHC class I expression on trophoblast, regulatory T-cell populations, and tryptophan-degrading IDO activity).
The fetus's own immune system matures gradually:
- Innate immunity is functional from early gestation, but neutrophil and complement function are reduced compared with adults.
- Adaptive immunity: lymphocytes are present from the first trimester, but antigen exposure is minimal, so naive cells dominate at birth.
- Maternal IgG crosses the placenta actively from around 20 weeks via the FcRn receptor, peaking in the third trimester. At term the fetus has IgG levels comparable to or slightly higher than its mother's. This passive immunity protects against many infections in the first 3-6 months until it wanes; the timing of routine infant immunisation reflects this.
- IgA does not cross the placenta but is delivered postnatally in breast milk, particularly the first colostrum, which is rich in secretory IgA.
Fetal CNS and Sensory Physiology
The fetal brain develops in a precisely timed sequence: neural tube closure by 4 weeks, neurogenesis from 8-25 weeks, neuronal migration through the second trimester, and synaptogenesis and myelination running from late gestation into early childhood. Disruption at any point produces a characteristic spectrum of malformations, set out in Development of the Nervous System.
By the late second trimester the fetus is responsive to sound, touch, light and pain. Hearing, in particular, is established sufficiently that maternal voice and rhythm are recognisable shortly after birth. Vision develops more slowly because of the dark intrauterine environment; visual acuity improves rapidly in the first months of life.
The fetal sleep-wake cycle, with periods of rapid eye movement (REM) sleep, is established by mid-gestation; non-REM sleep cycles emerge towards term. Fetal movements are detectable by the mother (quickening) at around 18-20 weeks in a primigravida.
Transition at Birth
Three coordinated changes happen in the seconds-to-minutes around birth:
- Lung aeration and breathing. The first breath generates very large negative intrathoracic pressures (sometimes −60 cmH2O) to overcome surface tension and inflate fluid-filled airways. With each subsequent breath, alveoli recruit and a functional residual capacity is established, helped by surfactant.
- Fall in pulmonary vascular resistance. Inflation, the rise in alveolar oxygen tension, and the drop in PCO2 all relax the pulmonary arterioles. Pulmonary blood flow increases roughly tenfold, and pulmonary arterial pressure falls.
- Closure of the fetal shunts:
- The foramen ovale closes functionally as left atrial pressure rises (now receiving the much larger pulmonary venous return) above right atrial pressure (now receiving less, because the placental return has stopped). Anatomical fusion takes weeks to months.
- The ductus arteriosus constricts in response to the rise in arterial oxygen tension and the fall in placental prostaglandins. Functional closure occurs within 24-48 hours; anatomical closure within a few weeks. The clinical importance of this prostaglandin-driven balance is exploited in opposite directions: prostaglandin synthesis inhibitors can close a persistently open ductus in the preterm neonate, while prostaglandin infusions can keep the ductus open in duct-dependent congenital heart disease until surgical correction. See Congenital Heart Defects.
- The ductus venosus closes within hours as umbilical blood flow stops. The umbilical vein and the distal portions of the umbilical arteries obliterate over the first weeks to leave the ligamentum teres (umbilical vein), the ligamentum venosum (ductus venosus) and the medial umbilical ligaments (distal umbilical arteries). The proximal segments of the umbilical arteries persist as the superior vesical arteries.
Two further neonatal challenges are worth noting:
- Thermoregulation. The neonate has a high surface-area-to-volume ratio and limited shivering. Heat is generated by non-shivering thermogenesis in brown adipose tissue, which expresses uncoupling protein-1 (UCP1) on its mitochondrial inner membrane. UCP1 dissipates the proton gradient generated by the electron transport chain as heat rather than ATP. Cold stress in a neonate burns through brown fat reserves, increases oxygen consumption, and can precipitate hypoglycaemia and acidosis; the rationale for warming the delivery room and skin-to-skin contact.
- Glucose homeostasis. Maternal glucose supply ends at delivery. Healthy term neonates draw on glycogen stores and start gluconeogenesis within the first hours, supported by the cortisol surge and falling insulin. Preterm and growth-restricted infants have smaller glycogen stores and a higher risk of hypoglycaemia; infants of diabetic mothers, with established hyperinsulinaemia, are at particular risk. Routine cord blood and early feeding regimens reflect this.
Summary
- The fetus exists in a low-oxygen, high-glucose environment supported by the placenta. Its physiology is built around maximising oxygen extraction and bypassing organs not yet needed.
- The fetal circulation has three shunts: the ductus venosus (a venous bypass of the liver, from umbilical vein to IVC) and two right-to-left cardiac shunts; the foramen ovale (atrial level) and the ductus arteriosus (great-vessel level): that bypass the pulmonary circulation.
- Fetal haemoglobin binds 2,3-BPG poorly, has a left-shifted oxygen dissociation curve, and is replaced by adult HbA over the first six months of life.
- Lung development proceeds through five overlapping stages; type II pneumocytes produce surfactant from around 24 weeks. Antenatal corticosteroids accelerate maturation in threatened preterm delivery.
- The fetal kidneys make amniotic fluid, not waste removal: oligohydramnios from renal disease causes pulmonary hypoplasia.
- The placenta transfers maternal IgG (passive immunity) but not IgA. Maternal hyperglycaemia drives fetal hyperinsulinaemia and neonatal hypoglycaemia.
- At birth the lungs aerate, pulmonary vascular resistance falls, and the three fetal shunts close. Brown adipose tissue drives non-shivering thermogenesis. Failure of any of these transitions underlies the major neonatal emergencies.
Reviewed by: Dr. Marcus Judge
In this article
The fetus exists in an oxygen-poor, glucose-rich, immunologically protected environment supplied by the placenta.
- 109

{kind=link}